
近日,中国科学技术大学生命科学学院及中科院分子卓越中心丁勇教授课题组与中国水稻所的钱前研究员和台湾大学的吴克强教授合作,发现组蛋白第4位赖氨酸三甲基化修饰 (H3K4me3) 调控开花时间和二级枝梗数的新机制,相关结果以“SIP1 participates in regulation of flowering time in rice by recruiting OsTrx1 to Ehd1”和“The COMPASS-like complex promotes flowering and panicle branching in rice”为题目,发表在4月3日的《New Phytologist》和近日的《Plant Physiology》杂志上。丁勇研究组长期从事植物组蛋白修饰调控机制的研究,这是继2017年《The Plant Cell》上三篇关于组蛋白修饰调控植物生长发育后,该研究组对组蛋白如何修饰调控目的靶基因的新发现。
DNA缠绕在组蛋白上,植物组蛋白甲基转移酶有结合DNA的结构域,又是如何特异结合靶基因,催化H3K4me3修饰,继而激活转录,并完成相应的生物学过程?研究表明,转录因子SIP1结合特异靶基因序列,招募甲基转移酶OsTrx1,完成靶基因的H3K4me3修饰。
水稻OsTrx1是TRX类蛋白,具有H3K4me3甲基转移酶活性。研究发现,OsTrx1与锌指蛋白SIP1相互作用,SIP1特异地结合开花靶基因Ehd1启动子15 bp的DNA序列,SIP1招募OsTrx1至Ehd1位点,完成H3K4me3修饰,并促进转录,完整地解释了功能缺失的SIP1和OsTrx1具有的早花表型。进一步研究发现,OsTrx1酶单独作用时甲基化效率低,OsTrx1甲基转移酶活性需要OsWDR5a辅助,OsWDR5a与OsTrx1相互作用后,提高OsTrx1催化H3K4me3修饰效率。在OsWDR5a功能缺陷条件下,即使OsTrx1结合Ehd1,也不能完成该位点H3K4me3修饰,解释了OsWDR5a功能缺陷,亦能导致与ostrx1类似的晚花表型。此外,功能下调控的OsTrx1和OsWDR5a不仅能调节水稻开花时间,而且影响二级枝梗数。该研究发现了组蛋白修饰甲基转移酶特异性修饰靶基因的分子机制,也为农作物性状改良提供分子基础。
丁勇课题组的客座博士研究生姜鹏飞为该工作的第一作者,其它作者还有同课题组的王世亮、郑撼等同学。该项研究在国家自然科学基金、中组部“青年创新人才计划”、中科院分子植物先导(B类)和中国科学技术大学微尺度国家研究中心共同资助下完成的。
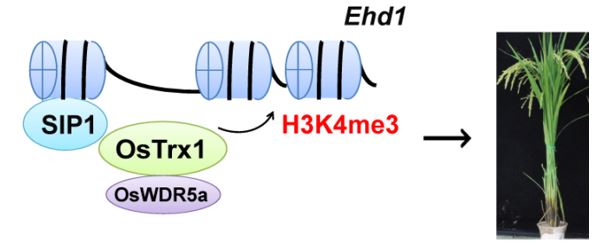
甲基化酶修饰靶基因的工作模式图
(生命学院、科研部)